

 POLSKI
POLSKI







Type 1 diabetes mellitus (T1DM) is a chronic autoimmune disease leading to immune-mediated destruction of pancreatic β cells, resulting in the need for insulin therapy. A genetic susceptibility to the development of islet autoimmunity and type 1 diabetes is well documented, and an environmental influence and impaired function of the immune system is assumed.
Genetic background
Polymorphisms of multiple genes of the human DNA can increase the risk of the disease. Approximately 50% of the genetic risk is associated with the HLA-DR and HLA-DQ HLA haplotypes, located in the major region of the tissue compatibility complex – MHC, which are responsible for antigen presentation to CD4 T lymphocytes by β cells, dendritic cells, and macrophages. HLA risk genes are responsible for the abnormal presentation of pancreatic islet antigens and the subsequent survival and expansion of β-cell reactive T cells, leading to pancreatic β-cell destruction. Most non-HLA genes are related to immune function and regulation (protein tyrosine phosphatase, PTPN22, IL2RA, CTLA4, Ins) [1].
Gene polymorphisms involved in sphingolipid metabolism and Vitamin D metabolism that contribute to the genetic predisposition to T1DM were also identified [2, 3].
Environmental factors
Environmental factors that increase the risk of developing T1DM include:
Viral infections (Coxsackie B virus, rubella viruses, enteroviruses, rotaviruses, CMV, retroviruses) through a mimicry mechanism [4].
Vitamin deficiency, mainly D and C through immunomodulatory effect [5, 6].
Factors that can affect the intestinal immune system, immune system, including diet (cow’s milk, gluten, toxins and chemical compounds, sweetened beverages) and hygiene. Some studies showed that early (< 4 months) and late (≥ 6 months) introduction of solid foods increased risk for development of isled antibodies in children at increased genetic risk, whereas breastfeeding had a protective effect [7, 8].
Changes in the gut microbiota. The intestinal flora is involved in the maturation of the innate and acquired immune system in early human life and in the formation of the intestinal barrier. The diversity of species depends on the type of birth, breastfeeding, and diet in early life. The use of antibiotics or improper nutrition lead to a disruption of the normal composition of the intestinal bacterium, the so-called dysbacteriosis, which, due to a decrease in the production of short-chain fatty acids (SCFAs) and loss of connections between enterocytes, increases intestinal permeability. Some bacteria show sequence similarities in the GAD of human gut bacteria and the GAD65 of the β-cell. Changes in the gut microbiome result in the release of bacterial GAD and the miseducation of antigen-presenting cells (APCs), which then recognise GAD65 pancreatic β cells [9–11].
Immune system
In genetically predisposed individuals, under the influence of environmental factors, β-cell autoantigens are recognised, subsequently presented by APCs (antigen-presenting cells). This leads to the activation of macrophages and T cells, which secrete pro-inflammatory cytokines that increase T cell (Thelper = CD4+ and Tcytotoxic = CD8+) and B cell activation. In addition, regulatory T cells have impaired number and function to suppress the activity and proliferation of CD4+ and CD8+ T cells. Eventually, effector cells (CD8+ lymph, NK cells, macrophages) and pro-inflammatory cytokines [mainly interleukin 1β (IL-1β), tumour necrosis factor α (TNF-α), and interferon γ (IFN-γ)] destroy the β-cell [12].
Stages of type 1 diabetes mellitus
The development of T1DM progresses sequentially at a variable but predictable rate through identifiable stages:
stage 1 is defined as the presence of β-cell autoimmunity, as shown by the presence of 2 or more anti-islet autoantibodies with normoglycaemia and no clinical symptoms,
stage 2 is defined as the presence of β-cell autoimmunity with dysglycaemia and without symptoms,
stage 3 is defined as the onset of symptomatic disease.
The adoption of the stage classification provides a standardised taxonomy of T1DM and helps in the development of therapies and the design of clinical trials to prevent the disease.
Antibodies produced by B cells are useful to determine the diagnosis and stage of CT1 development, as well as in prevention trials [13].
It has been estimated that children with multiple β cell autoantibodies have a 70% risk of developing type 1 diabetes in 10 years and a lifetime risk approaching 100%.
The main pancreatic islet autoantibodies produced by B lymphocytes are insulin autoantibodies (IAA), insulinoma-associated antigen-2 (IA-2), glutamic acid decarboxylase (GAD), Zn transporter 8 (ZnT8), and pancreatic islet cell antibodies (ICA). These autoantibodies can appear at any age, but the peak of the first pancreatic islet autoantibody usually occurs before the age of 3 years [14].
Most autoantibodies associated with T1DM recognise peptide antigens, but antibodies against lipid antigens (p/gangliosides and sulfatides) have also been described [15, 16].
Prevention of type 1 diabetes mellitus
Prevention of T1DM is divided into the following (Fig. 1):
Primary prevention, which applies to people at risk of developing T1DM and includes actions aimed at preventing the onset of autoimmunity by addressing risk factors.
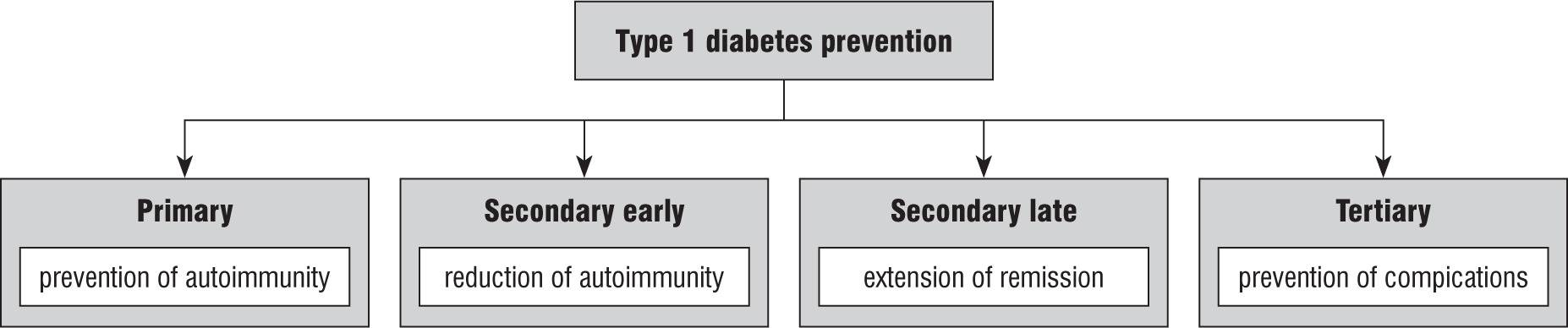
Secondary prevention – the aim is to inhibit or slow down autoimmune reactions and delay symptomatic CT1 or prolong the period of remission. Secondary prevention is divided into early pre-clinical, which is undertaken when autoimmunity has already started but the number of β-cells is sufficient to maintain glucose homeostasis, and late secondary prevention in people who have developed impaired glucose tolerance, or in the early period after the onset of type 1 diabetes.
Tertiary prevention, which aims to prevent complications of the disease.
Environmental Interventions
Initial results from DAISY showed an association between enterovirus infection and acceleration in disease onset in high-risk individuals, which suggests an enterovirus vaccine serving as a potential primary or secondary prevention therapy for T1DM [17].
Human studies of the gut microbiome in T1DM have shown that early probiotic supplementation (at the age of 0–27 days) was associated with a decreased risk of islet autoimmunity among children at increased genetic risk of T1DM [18].
The GPPAD-SINT1A Study assessed whether early Bifidobacterium infantis supplementation reduces the risk of islet autoimmunity in children at the highest genetic risk of T1DM [19].
In secondary prevention, the use of probiotics/synbiotics was associated with reduction of pro-inflammatory cytokines (IL-1, IL-8, IL-17, MIP-1β, RANTES, and TNF-α), oxidative stress (hs-CRP), and an increase in total antioxidant capacity and the anti-inflammatory cytokine TGF-β1. Meta-analyses evaluating the effect of probiotics showed HbA1c and fasting blood glucose reduction [20–22].
Immunotherapy
Historical immunotherapy (such as cyclosporine, islet transplantation with chronic immunosuppression) highlighted the fact that altering the immune system needs to be done in a balanced way that preserves safety with efficacy [23, 24].
Designing successful immune interventions for autoimmune diseases like T1DM relies on understanding the interactions between genes, the environment, and immune regulation.
Targeting of immunotherapy towards the islet-specific immune pathways has led to the development of therapies that may allow for the prevention or reversal of this disease while avoiding toxicities.
Antigen nonspecific therapy
T lymphocytes are the final element leading to the destruction of β-cells. CD3, CD4, and CD8 antigens are presented on T lymphocytes, so therapies targeting these antigens can modulate the T-cell response and therefore modulate the autoimmune response.
Targeted CD3 blockade therapies by teplizumab, a humanized monoclonal antibody to CD3 on T cells, is approved by the US Food and Drug Administration to delay the onset of clinical type 1 diabetes (stage 3) in patients 8 years of age or older with preclinical (stage 2) disease. In children and adolescents with newly diagnosed type 1 diabetes, it showed benefit with preservation of β-cell function [25, 26].
Low-dose ATG is a purified rabbit sera containing cytotoxic IgG antibodies directed against human T cells, which was able to slow the rate of β-cell function and reduce HbA1c [27].
B cells also have a pathogenic role in T1D as antigen-presenting cells and modulators of the pancreatic microenvironment. Immunotherapy with the anti-CD20 monoclonal antibody rituximab against B lymphocytes slightly delays the decline of C-peptide [28].
Immunotherapies with rituximab, abatacept, teplizumab, alefacept, high-dose anti-thymocyte globulin (ATG), low-dose ATG, and low-dose ATG ± granulocyte-colony-stimulating factor have demonstrated endogenous insulin preservation in recent-onset T1DM. Low-dose ATG and teplizumab show the greatest impact on C-peptide preservation among recent new-onset T1DM studies [29].
Antigen-specific therapy
Antigen-specific intervention is a promising approach that may avoid the side effects seen with systemic immunomodulating therapies. According to preclinical models, single antigen-specific therapies are capable of suppressing the polyclonal immune response. The exposure to peptides of T1DM specific auto-antigens (e.g. β cell auto-antigens) can induce an expansion of the Tregs and the deletion and/or anergy of pathogenic T cells, thereby restoring immune tolerance.
Immunization with insulin, proinsulin, GAD65, GAD-alum, and a mixture of peptides from islet autoantigens (MultiPepT1De) has not resulted in preserved insulin secretion so far, but some of the clinical trials are still ongoing [30].
The autologous ex vivo-expanded polyclonal Tregs were well tolerated and safe but unable to preserve C-peptide [31].
Cellular approaches (i.e. Tregs) may be the most attractive approach to restore immune tolerance because they bypass unknown autoantigens and to who should be introduced antigen in a true tolerogenic form.
Targeting inflammation
Whether therapies targeting the generalized inflammatory response with blockade of IL-1 using anakinra and canakinumab, the pro-inflammatory cytokine tumour necrosis factor (TNF) using etanercept, golimumab, anti-IL-6 (tocilizumab), or anti-IL-21 by liraglutide can restore immune tolerance in T1D [32, 33] remains to be seen.
Therapies focused on maintaining β cell mass
Recent evidence has demonstrated that, among other factors, reduced amounts of sulfatide may be relevant in the development of T1DM. It stimulates calcium ion-dependent insulin exocytosis, promotes proinsulin folding, mediates monomerization of insulin, induces glucose-dependent stimulation of insulin, and prevents apoptosis in insulin secreting cells. Fenofibrate, which activates sulfatide biosynthesis, is evaluated in the PRIFEN Study to ascertain if it can be a target for improving outcomes in subjects with T1DM [34–36].
Combining immunotherapies target different mechanisms of T1DM development represents a way to optimally suppress pathogenic pathways and stimulate regulatory pathways that may result in immune tolerance restoration.
The effectiveness of immune therapies also depends on the selection of the patient group.
Mechanisms leading to T1DM differ in different groups; usually the autoimmune process is more aggressive in children.
Biomarkers reflecting disease activity and patient status can predict who will respond to a particular immunotherapy.